Organ Specificity
In a previous study on U. maydis mutants with deletions for clusters encoding secreted effectors, more than 50% of the mutants showed no phenotype or only marginal virulence defects (Kämper et al. 2006, Nature). This raised the question, why U. maydis has retained these effector genes. In this context, we hypothesized that those effectors might allow U. maydis to tailor its weaponry to the specific conditions characteristic for different host organs. Parallel transcriptome profiling of both U. maydis and Z. mays genes in seedlings, adult leaves and tassels, validated this hypothesis and revealed dramatic differences in gene expression of both the host and the pathogen in the different organs (Skibbe et al., 2010, Science). Using a gene deletion strategy, we could demonstrate that individual effector genes contribute to U. maydis virulence in an organ-specific manner (Schilling et al., 2014, Molecular Plant Pathology). One effector that specifically contributes to tumor formation in maize leaves is See1 (Seedling efficient effector1). This effector is translocated to the host cell, where it interacts with the maize SGT1 protein, a central regulator of cell-cycle regulation and immunity. Strikingly, See1 is required to establish U. maydis-induced cell cycle progression and therefore actively involved in triggering plant tumors (Redkar et al., 2015, Plant Cell).
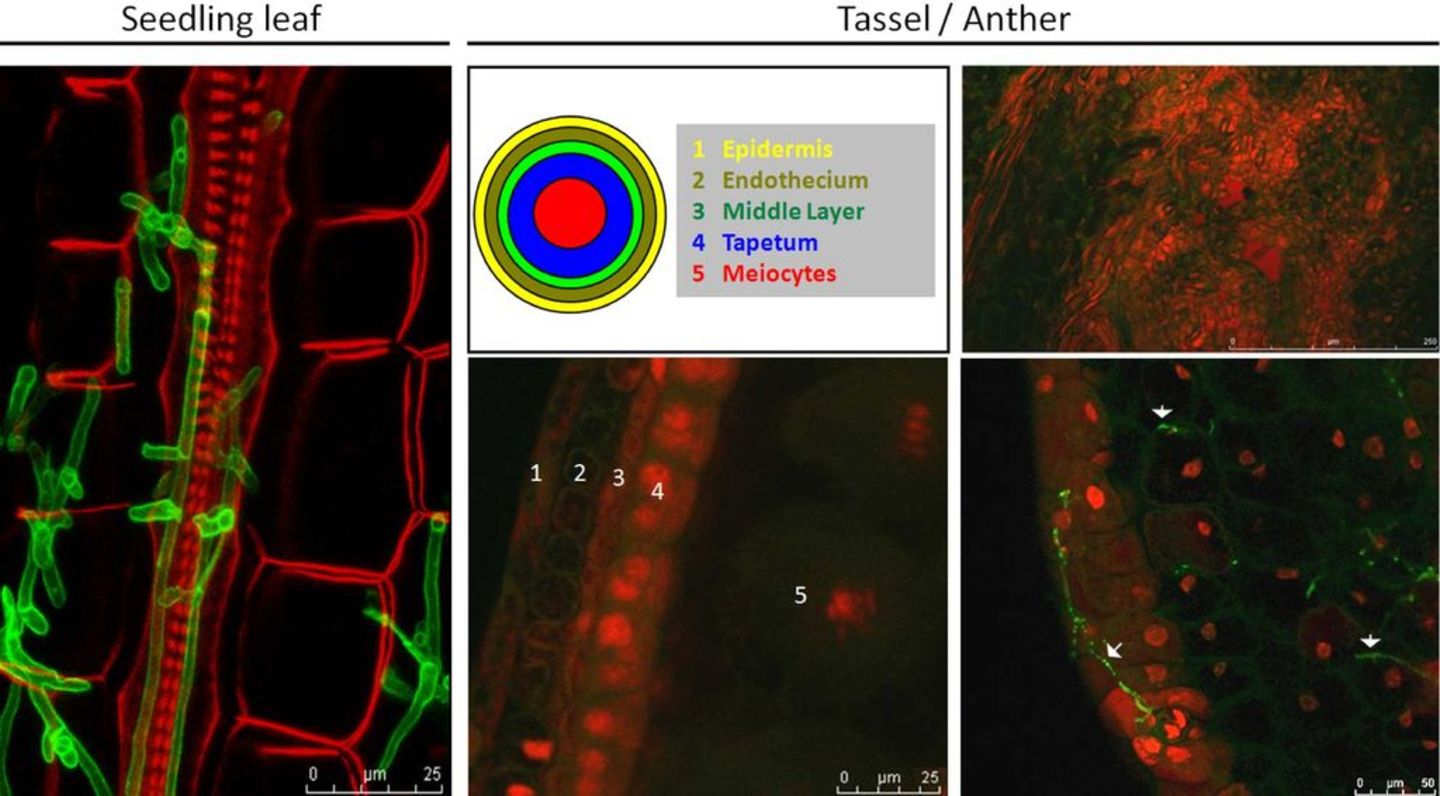
Figure 2: Biotrophic development of U. maydis in seedling leaf tissue and anthers. Left panel: Biotrophic U. maydis colonizing a seedling leaf (4 dpi). Fungal hyphae (WGA-AF488: green) grow both intra- and intercellularly, mostly around vascular bundles that are indicated by strong cell wall fluorescence (red). Middle panel: Maize anthers are four lobes surrounding the connective tissue; centrally there is a single vascular strand in the middle of the connective tissue. Architecture of maize anther locules with 5 distinct cell layers (numbered 1-5) as indicated schematically and in a confocal projection. Red: propidium iodide staining of plant DNA; grey: plant cell walls. C: U. maydis infected maize anther 8 dpi. Right panel: The defined structure of the anther is completely obscured, the anther tumor consists of undifferentiated, densely packed plant cells (upper right image). In the anther tumor cells, fungal hyphae (WGA-AF488: green) grow intracellularly (lower right image, arrows).
We are currently investigating the organ-specific functions of the group of organ-specific effector proteins to elucidate how U. maydis adapts to different host environments and deploys specific effector proteins that comprise cell-type specific host targets. Moreover, we aim to move on from the organ- to the cell type-specific level, following our hypothesis, that effectors act on individual cell types to guide transition into tumors (Figure 3).
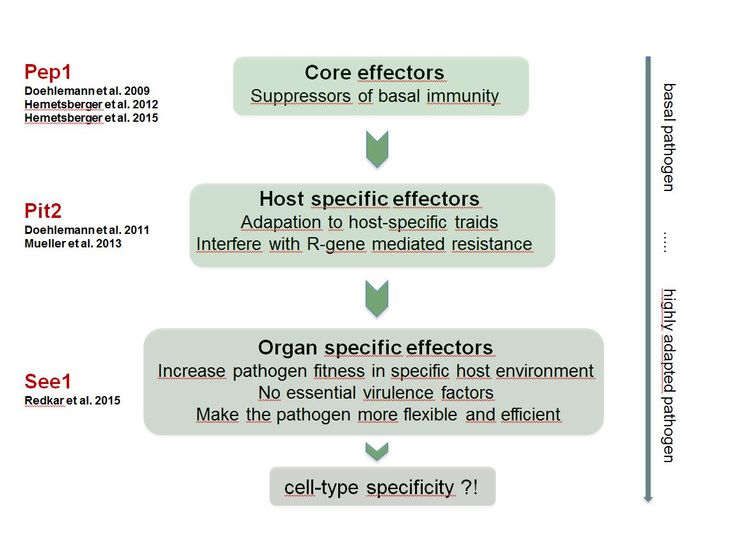
Figure 3: Model of effector specialization during co-evolution of host and pathogen