Molecular Functions of Effector Proteins
Plant colonizing microbes secrete a plethora of molecules to the host tissue to facilitate infection. Such molecules that interfere with host processes to the favor of the pathogen are generally called “effectors”. Our group is mainly interested in proteins/peptides (=> effector proteins) that are secreted by the pathogen. The research strategy is based on a reverse genetics approach; candidate genes encoding for putative effector proteins are deleted in the fungus and the resulting mutants are tested for alterations in virulence. This approach already identified core effectors of U. maydis, which hold crucial functions for virulence.
The secreted effector protein Pep1 is specifically expressed during pathogenic development of U. maydis. Deletion mutants for pep1 show no defect during saprophytic growth, but are arrested upon host penetration and elicit a hypersensitive response of the plant (Doehlemann et al., 2009, PloS Pathogens; Figure 1).
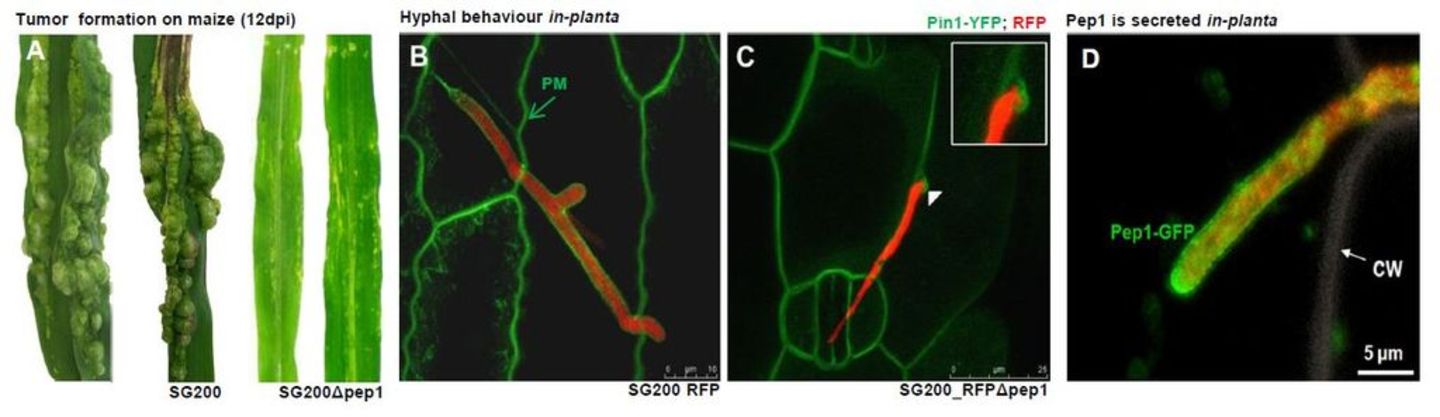
Figure 1: The Ustilago maydis effector Pep1. A: Phenotype of virulent U. maydis strain SG200 compared to the pep1 deletion mutant SG200Δpep1, both 12 days after infection. B,C: SG200 and SG200Δpep1 strains expressing cytoplasmic RFP 24 hours after infection on maize plants expressing the plasma membrane marker PIN1-YFP (Doehlemann et al., 2009). SG200 colonizes living maize cells and is surrounded by the plasma membrane, while the pep1 deletion mutant stops directly after invagination of the host membrane. D: Pep1-GFP is secreted from a hyphal tip that is growing inside a maize epidermis cell. CW: plant cell wall.
Thus, a biotrophic interaction is not established and the infected plant tissue shows various defense responses, particularly the production of reactive oxygen species (ROS). We identified Pep1 as an efficient inhibitor of the plant oxidative burst response, which is elicited by so called pathogen associated molecular patterns (PAMPs). This inhibitory effect of Pep1 on the early plant defense is explained by an inhibition of plant peroxidase activity through Pep1 (Hemetsberger et al., 2012, PloS Pathogens). Suppression of this conserved component of the plant ROS generating system by a pathogen effector therefore shows a new strategy of biotrophic pathogens to suppress innate plant immunity. A striking finding for Pep1 was its strong conservation in other smut fungi, including the dicot pathogen Melanopsichium pennsylvanicum. This conservation is not only on sequence level but also functional, as all orthologs tested were able to complement virulence in the U. maydis pep1 deletion mutant (Hemetsberger et al., 2015, New Phytologist).
Another secreted effector of U. maydis is Pit2. The pit2 gene is located in a small cluster of four genes (the pit-cluster) of which two encode proteins being essential for U. maydis virulence, the membrane protein Pit1 and the secreted effector Pit2 (Doehlemann et al., 2011, Mol Microbiol). Similarly to Pep1, Pit2 is specifically produced during plant colonization and it is also secreted to the biotrophic interface. However, while pep1 mutants are blocked by early plant defenses, pit2 deletion mutants are able to establish a biotrophic interaction but fail to induce tumor formation and elicit host defense at later steps of infection. Pit2 was found to inhibit a group of apoplastic plant proteases (papain like cysteine proteases; PLCPs), which are important factors of plant immunity (Mueller et al., 2013, PloS Pathogens). More details on this topic you can find here.
Since effector proteins that serve as virulence factors are precious molecular tools to identify key components of the plant immune system, we are continuously interested in the functional characterization of novel effectors.